



















 수량을 선택해주세요.
수량을 선택해주세요.




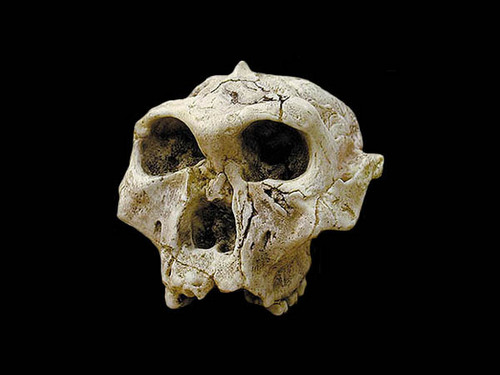
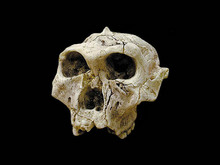
일반명 : 오스트랄로피테쿠스 로부스투스
학명: Australopithecus robustus (SK-48)
시대 : 1.5~2 million-year-old
발견지역 : Fourie in Swartkrans, South Africa
발견시기 : 1950
발견자 : Robert Broom in 1952.
1.5 to 2 MYA. The Australopithecus robustus Skull SK-48 was discovered by Fourie in
Written by C. David Kreger
Introduction
The species Australopithecus robustus was first discovered and named by the eminent Dr. Robert Broom. Broom made a habit of buying fossil remains from a lime quarry worker, and on a particular visit on June 8, 1938, Broom bought a maxillary fragment containing a first molar. The shape and the size of the molar convinced Broom that this was a different species than A. africanus (Broom's transvaalensis), and upon further investigation, found that the specimen had been found by a young boy who worked in the cave as a guide on Sundays. Broom searched for the boy (Gert Terblanche) and found him at school. Broom lectured the boy's class on the cave sites of the area, and was then led to the place of the specimen's discovery, Kromdraai. Broom found several more cranial and mandibular fragments associated with the original maxillary specimen, and this partial cranium (TM 1517) became the type specimen for A. robustus.
Broom spent some time working on a monograph of the australopithecines, which was published in 1946. This monograph included the description of TM 1517, and was a turning point for the South African australopithecines in the eyes of the world. The monograph received the U.S. National Academy of Sciences award for the most important book of the year in biology, and along with L Gros Clark's published approval of the South African australopithecines as hominids, was very important in altering the view that the South African specimens were human ancestors and not simply an ape.
Another important find was SK 6, a portion of mandible and several teeth. Another discovery of Broom's, he originally named a new species for it (Paranthropus crassidens.) However, the specimen is generally accepted as robustus, though it is also considered a type specimen because of this separate species designation. One of the more complete robustus specimens is SK 48, a fairly complete crania of what is probably a female. This specimen - due to its completeness - added much information on the cranial characteristics of the robust australopithecines. One specimen may or may not be robustus, since it was found at Sterkfontein, and the presence of robustus and africanus in the same place at basically the same time would cause some phylogentic attribution of the robust australopithecines major problems. STW 252 is considered by those who see more than one species at Sterkfontein as robustus, and as africanus by those that see one species. The dentition seems to indicate inclusion in robustus, but the uncertainty of the date causes some to view it cautiously.
Diagnostic Features
The A. robustus remains generally are from three sites: Swartkrans, Dreimulen, and Kromdraai (with the aforementioned Sterkfontein specimen possibly included.) By far the largest of these sites is Swartkrans. One of the major problems with these South African sites is dating, but generally, robustus remains can be safely placed from 2.0-1.0 myr, and possibly even earlier. The dating of these sites is crucial to understanding the phylogeny of the robust australopithecines, but for now, the dates are somewhat in question. An understanding of the characteristics of robustus can best be seen by comparing them to an earlier specie (africanus) and a penecontemporary one (boisei).
The robustus crania are many, but fragmentary, with a known cranial capacity for just one individual specimen, SK 1585, an endocast with a 530 cc capacity. There is evidence of significant expansion over africanus, with an estimated 15% average increase in brain size over africanus. The sexual dimorphism level of about 20% seems to be basically unchanged. In his analysis of SK 1585, R. Holloway concluded that robustus shows a general trend towards a more modern brain - similar to that of boisei - over that of africanus. These modern trends include:
- Some structural brain differences reflected in endocasts, such as: A) greater frontal lobe breadth B) expanded parietal cortex C) increased cerebral height (high cerebral to cerebellar height) D) cerebellar lobes "tucked in" and not projecting laterally or posteriorly.
- Increased flexion of the cranial base.
- Shortening of the base and decrease in the angle of the petrous pyramids.
- More anterior foramen magnum position.
- Deeper mandibular fossa with well-delineated, projecting, articular eminence.
- Nearly horizontal orientation of nuchal plane.
- Expanded height of occipital plane of the occiput, with a concomitant low inion position.
- Decreased facial prognathism, especially subnasal.
- Shortened distance between the tooth row and the mandibular fossa.
- Reduced posterior component of temporalis muscle.
- Weakly developed or absence of pneumatized bone in the temporal squama.
The increase in body size over africanus is minimal, which means that there is significantly increased encephalization, rather than simply an allometric increase with increased body size. The remains from Swartkrans are very variable, but seem (compared to africanus) to have more robust crania, with better developed muscle markings, more prominent tori, and thicker buttressing structures. Like the earlier specimens, these features are on a fairly thin cranial vault.
The cranial bases seem to differ, but the preservation makes comparisons difficult, with only one unambiguously preserved specimen (SK 47). What evidence there is seems to indicate that the Swartkrans hominids have a broader and shorter (more Homo-like) cranial bases with a more anterior foramen magnum position, and a petrous bone that is more transversely oriented than in earlier australopithecines. The amount of basicranial flexion seems moderate (more than africanus, less than boisei). However, these observations are made unclear due to the specimen being a juvenile, but the 495 cc cranial capacity is close to the female mean, and there likely would have been little change.
A. robustus continues the trends seen in africanus with regards to facial changes, but remains very similar (yet distinct) from boisei). These features include:
- The two species are equally prognathic.
- Both have nasal bones that are the same size and shape.
- Both have anterior pillars that border the nasal aperture, extending upward from the buttresses for the canine roots.
- Both have a lower border of the cheek that is virtually a straight line from its origin on the side of the palate, extending to the base of the zygomatic arch, which is the widest portion of the face.
- There is substantial increases in postcanine tooth size (though not at the level of boisei), with increases in both absolute and relative size over africanus.
The canines seem to have changed little, while the incisors decreased in size significantly, possibly due to overcrowding, as there is overlap of anterior teeth in 43% of the Swartkrans remains. The increase in postcanine tooth size includes an increase in enamel thickness, and while this has been interpreted in various ways, the idea that robustus was an herbivore/frugivore that subsisted on hard gritty nuts and plants (J.T. Robinson's dietary hypothesis) seems to be in serious doubt, with an eclectic omnivorous diet seems to be more likely due to recent work determining low strontium/calcium ratios, and C4 contribution (likely from eating grazing animals).
Postcranial evidence shows little difference between robustus, africanus, or boisei. They were all small-bodied obligate bipeds, that used an efficient walking gait.
Conclusions
The main question regarding A. robustus seems to be the question of whether or not it is a separate species from A. boisei, or if they are geographic species of a wide-ranging variable population. Most researchers seem to agree that they are separate lineages, with several important evolutionary trends that distinguish them. However, they share even more of the same evolutionary trends. It seems safe to call these separate lineages, but a vocal protest exists among those who see a monophyletic lineage (re: Paranthropus).
Bibliography
Aiello, L., and C. Dean. 1990. An Introduction to Human Evolutionary Anatomy.
Broom, R.A. 1938. "The Pleistocene anthropoid apes of
Broom, R.A. 1949. "Another new type of fossil ape-man." In Nature, vol. 163, pg. 57.
Broom, R.A., and J.T. Robinson. 1950. "Man contemporaneous with the Swartkrans ape-man." In American Journal of Physical Anthropology, vol. 8, no. 2, pp. 151-155.
Broom, R.A., and J.T. Robinson. 1952. "Swartkrans ape-man, Paranthropus crassidens." In
Holloway, R.L. 1988. "'Robust' australopithecine brain endocasts: Some preliminary observations." In Evolutionary History of the "Robust" Australopithecine, ed. by F.E. Grine, pp. 97-105.
Johanson, D., and B. Edgar. 1996. From Lucy to Language.
Jones, S., R. Martin, and D. Pilbeam, eds. 1992. The Cambridge Encyclopedia of Human Evolution.
Lee-Thorpe, J.A., N.J. van der Merwe, and C.K. Brain. 1994. "Diet of Australopithecus robustus at Swartkrans from stable carbon isotope analysis." In Journal of Human Evolution, vol. 27, no. 4, pp. 361-372.
McHenry, H. 1994. "Behavioral ecological implications of early hominid body size." In Journal of Human Evolution, vol. 27, pp. 77-87.
Rak, Y. 1983. The Australopithecine face.
Robinson, J.T. 1956. "The dentition of the Australopitho-pithecinae." In
Sillen, A. 1992. "Strontium-calcium ratios (Sr/Ca) of Australopithecus robustus and associated fauna from Swartkrans." In Journal of Human Evolution, vol. 23, no. 6, pp. 495-516.
Sillen, A. 1996. "Physiological variability and Sr/Ca at Swartkrans: A comment on Thackeray, 1995." In Journal of Human Evolution, vol. 30, no. 4, pp. 381-384.
Wolpoff, M. 1999. Paleoanthropology. second edition.