




















 수량을 선택해주세요.
수량을 선택해주세요.




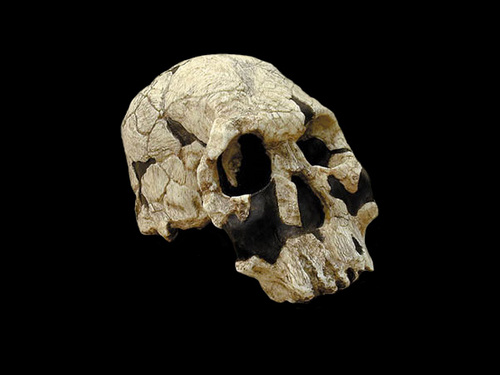
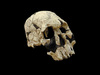
일반명 : 호모 루돌펜시스
학 명 : Homo rudolfensis (KNM-ER 1470)
시 대 : 1.8 to 1.9 MYA.
발견지역 : Koobi Fora in Kenya
발견시기 : 1972
발견자 : B. Ngeneo
발표 : described by R. Leakey in Nature in 1973.
1.8 to 1.9 MYA. The Homo rudolfensis Skull KNM-ER 1470 was discovered by B. Ngeneo in 1972 at Koobi Fora in
Written by C. David Kreger
Introduction
The species designation of Homo rudolfensis is a much debated topic, over both whether it is a separate species, and if it is an australopithecine rather than a member of the genus Homo. The type specimen of the species is KNM-ER 1470. This specimen was discovered by Richard Leakey's team in 1972, east of Lake Turkana at Koobi Fora in northern
In 1986, Russian anthropologist Valerii Alexeev applied the species name of Pithecanthropus rudolfensis to ER 1470. Despite the incompleteness of the description, and Alexeev's unfamiliarity with the original specimen, the name stuck. The genus name of Pithecanthropus has been dropped by those who see rudolfensis as a valid species and replaced with the genus Homo. Recently, there have been attributions of the rudolfensis specimens as belonging to Australopithecus, and thus another genus change may be in the future of this species if it retains validity as a separate species.
One of the main problems with the rudolfensis species is that there are no postcranial remains that are associated with cranial remains. The rudolfensis specimens have large brains in conjunction with megadont postcanines, and without postcranial evidence it is unknown whether these features are due to a larger body size than contemporary habilis specimens. Due to this problem, competing ideas abound regarding the validity of rudolfensis and its proper place in hominid phylogeny. Some researchers see the larger brain and tooth size as indicative of allometric changes due to increased body size, with rudolfensis and habilis constituting the same species, with the former the males and the later the females. Some see rudolfensis as the ancestor of habilis with a decrease in brain size occurring, and others see the two on completely different evolutionary lines.
This debate is long from over, and all the scenarios have one problem or another. Perhaps with future discoveries that are attributed to the rudolfensis, a clearer picture will appear as to the relationship of these early Homo.
Diagnostic Features
The type specimen of Homo rudolfensis is KNM-ER 1470, discovered by Bernard Ngeneo at Koobi Fora,
Compared to a smaller Koobi Fora cranium, KNM-ER 1813, ER 1470 shows many differences that distinguish the specimen from habilis. These features include:
- A slight supraorbital torus across the forehead with no sulcus or depression behind it.
- A much longer face, with the upper part narrower than the middle.
- The maxilla is squared off rather than rounded, with a very short, shallow palate.
- Evidence of much more megadont postcanines.
Other features that comprise ER 1470 (but which are not necessarily distinct to the species) include:
- A lack of crests and heavy muscle markings that characterize australopithecine crania.
- A marked constriction of the braincase behind the orbitals (but less that occurs in robust australopithecines. )
- A bulging frontal bone that rises steeply to meet the square parietal bone that form the thin-walled sides of the brain case.
- An occipital bone that is smoothly rounded rather than flexed as in Homo erectus.
Other cranial remains that have been attributed to rudolfensis include KNM-ER 1590, KNM-ER 3732, KNM-ER 1801, and KNM-ER 1802. Alexeev originally attributed ER 1470 as a male rudolfensis and ER 1813 as a female, but ER 1813 is generally considered to be habilis. There is no directly associated postcrania for rudolfensis (which makes the species problematic), though some specimens have been attributed to the species. Limbs such as the ER 1472 and ER 1481 femora have been attributed to rudolfensis, suggesting a dramatic allometric difference, but there is much questionable about associating postcrania and cranial material together, when there is no objective sample to compare them to (an associated cranium and postcranial material).
Another specimen that may be attributable to rudolfensis is OMO 75-14, a mandible and cranial fragments that have been dated to around 2.0 myr. In spite of very large postcanine size (the M2 size exceeds all other non-robust specimens and its shape resembles the boisei condition), its premolar morphology is not like any robust australopithecine, according to G. Suwa et al. The teeth are larger than any habilis specimen, and seem to show definite Homo-like patterns. However, isolated mandibles and fragments are hard enough to attribute, and even worse when trying to attribute them to a species many researchers do not even consider a valid taxon. B. Wood has given a diagnosis of mandibles that should be placed within rudolfensis which include:
- Broader postcanine tooth crowns; they are narrower in habilis.
- A large P4 talonid (an extra area on the back of the tooth that acts to make it squarer in profile).
- Double-rooted P4s and sometimes P3s (these are single-rooted in habilis).
The dating of this species is significant, in that a date earlier than habilis makes this species the first habiline, and with its very large brain, a candidate for being a direct human ancestor. The initial date of 2.9 myr for ER 1470 has been modified, but as mentioned above, there is some support for a date of approximately 2.4 myr for the specimen. However, habilis itself has had its origins pushed back to approximately 2.3 myr (A.L. 666-1), making the designation of the "first" habiline difficult even if ER 1470 is older than is now generally accepted. Also important when considering the habilines (and rudolfensis in particular) is the idea that they are not Homo at all, but rather Australopithecus.
Analyses by D. Lieberman and B. Wood seem to indicate that the habilines share more features with the australopithecines than with Homo. Evidence for this position include:
- A. Walker's initial impression of ER 1470 was of an expanded but very similar specimen to STS 5, an africanus specimen.
- D. Lieberman et al. analysis which showed that rudolfensis shows more affinities to the australopithecines rather than with the other species of Homo.
- It would help account for the robust australopithecine features found in rudolfensis, because africanus closely resembles the robusts in many ways.
The large degree of variation in africanus shows that the degree of difference between rudolfensis and earlier africanus is such that attribution to differing species is not required.
Conclusions
As stated, the attribution of the species rudolfensis to any specimen is somewheat controversial, since many paleoanthropologists do not see rudolfensis as a valid species. Its dating (whether the early dates proposed by some or the contemporaneous dates to habilis) makes its brain size an issue, and raises questions about current standard phylogeny of the human line. Homo rudolfensis may be the first member of the genus Homo on a path to modern humans, or it may be a more Homo-like australopithecine with no direct bearing on the evolution of H. sapiens. Nothing can be stated for sure at this point, except that there will be much more future debate on the issue.
Bibliography
Alexeev, V.P. 1986. The Origin of the Human Race.
Grine, F.E., W.L. Jungers, and J. Schulz. 1996. "Phenetic affinities among early Homo crania from East and
Johanson, D., and B. Edgar. 1996. From Lucy to Language.
Kramer, A., S.M. Donnelly, J.H. Kidder, S.D. Ousley, and S.M. Olah. 1995. "Craniometric variation in large-bodied hominids: Testing the single-species hypothesis for Homo habilis." In Journal of Human Evolution, vol. 29, no. 5, pp. 443-462.
Leakey, R.E. 1973. "Evidence for an advanced Plio-Pleistocene hominid from East
Leakey, R.E. 1976. "An overview of the Hominidae from East
Lieberman, D.E., D.R. Pilbeam, and B.A. Wood. 1988. "A probablistic approach to the problem of sexual dimorphism in Homo habilis: A comparison of KNM-ER 1470 and KNM-ER 1813." In Journal of Human Evolution, vol. 17, no. 5, pp. 503-511.
Lieberman, D.E., B.A. Wood, and D.R. Pilbeam. 1996. "Homoplasy and early Homo: An analysis of the evolutionary relationships of Homo habilis sensu stricto and H. rudolfensis" In Journal of Human Evolution, vol. 30, no. 2, pp. 97-120.
Miller, J.M.A. 2000. "Craniofacial variation in Homo habilis: An analysis of the evidence for multiple species." In American Journal of Physical Anthropology, vol. 112, no. 1, pp. 103-128.
Rak, Y. 1985. "Systematic and functional implications of the facial morphology of Australopithecus and early Homo." In Ancestors: The Hard Evidence, ed. by E. Delson, pp. 168-170.
Rightmire, G.P. 1993. "Variation among early Homo crania from
Strait, D.S., F.E. Grine, and M.A. Moniz. 1997. "A reappraisal of early hominid phylogeny." In Journal of Human Evolution, vol. 32, no. 1, pp. 17-82.
Stringer, C. 1986. "The credibility of Homo habilis." In Major Trends in Primate and Human Evolution, ed. by. B. Wood, L. Martin, and P. Andrews, pp. 266-294.
Stringer, C. 1987. "A numerical cladistic analysis for the genus Homo." In Journal of Human Evolution, vol. 16, no. 1, pp. 135-146.
Stringer, C. 1992. "Homo habilis closely examined." In Current Anthropology, vol. 33, no. 3, pp. 338-340.
Tattersall,
Wolpoff, M. 1999. Paleoanthropology. second edition.
Wood, B.A. 1991. Koobi Fora Research Project (volume 4). Hominid Cranial Remains.